Rozmnožovanie rastlín je základným procesom, ktorý zabezpečuje zachovanie druhu a prenos dedičných vlastností z rodičov na potomkov. Tento fascinujúci proces môžeme rozdeliť do dvoch hlavných skupín: nepohlavné (vegetatívne) a pohlavné rozmnožovanie.
Nepohlavné (vegetatívne) rozmnožovanie
Nepohlavné rozmnožovanie vyžaduje len jeden materský organizmus. V tomto prípade vzniká nový jedinec z vegetatívnej časti materského organizmu, pričom jeho genotyp je zhodný s materskou rastlinou. Bunkové delenia prebiehajú výlučne mitózou, vďaka čomu vznikajú geneticky identické klony. Ide o energeticky výhodnú stratégiu na rýchlu kolonizáciu stabilného prostredia bez závislosti od opeľovačov. Nepohlavné rozmnožovanie je tiež kľúčové pre druhy s poruchami meiózy, napríklad pri zmnožení chromozómových sád (polyploidia).
Jednoduchšie organizmy využívajú na rozmnožovanie základné fyziologické formy:
- Bunkové delenie - typické pre sinice a jednobunkové riasy.
- Fragmentácia stielky - rozpad stielky na životaschopné časti. Využívajú ju napríklad mnohobunkové sladkovodné spájavky, ktorých vlákno po roztrhnutí jednoducho dorastie na novú riasu.
- Výtrusy - jednobunkové útvary na šírenie, u nižších húb a organizmov tvorené aj mitoticky.
Zložitejšiu stratégiu na nepohlavné šírenie majú niektoré machorasty. Nový jedinec môže vznikať zo spóry (výtrusu), ktorá sa tvorí v sporangiofóroch (výtrusniciach) = výtrusné rastliny (machorasty, prasličky, plavúne, paprade). Výtrusy sú nepohlavné rozmnožovacie častice, ktoré sú výsledkom redukčného delenia.
Vyššie rastliny si na vegetatívne rozmnožovanie vyvinuli špecializované orgány vzniknuté premenou stonky, koreňa či listov:
- Rozmnožovacia cibuľka - pozostáva z podcibulia a zdužnatých zásobných listov. Tento typ množenia je charakteristický pre čeľaď ľaliovité (napríklad obľúbený tulipán alebo cesnak, ktorý vytvára zložené dcérske cibuľky) a skorú jarnú čeľaď amarylkovité (snežienka, narcis).
- Podzemok (rizóm) - je vodorovne rastúca podzemná stonka. Z jej uzlov rastú korene a púčiky formujúce nové rastliny. Je to bežný jav predovšetkým u papradí, z kvitnúcich rastlín ho využíva napríklad liečivý kostihoj alebo mimoriadne odolný burinný pýr.
- Podzemková hľuza - je zhrubnutý zásobný podzemok stonkového pôvodu. Najlepším príkladom je zemiak, ktorého známe „očká“ sú v skutočnosti pazušné púčiky novej rastliny.
- Stonková hľuza - je nadzemná alebo podzemná metamorfóza plniaca zásobnú aj reprodukčnú funkciu. Typickým zástupcom s nadzemnou hľuzou je kaleráb. Veľmi podobný mechanizmus, avšak vo forme koreňovej hľuzy, si vytvorila reďkovka.
- Poplaz (stolón) - ide o horizontálnu plazivú stonku, ktorá rastie po povrchu a zakoreňuje priamo vo svojich uzloch. Tento efektívny mechanizmus plošného šírenia má jahoda.
- Listy - na nepohlavné rozmnožovanie môžu slúžiť priamo aj listy.
Pohlavné rozmnožovanie
Pohlavné rozmnožovanie spája genetický materiál dvoch jedincov, čím zabezpečuje nevyhnutnú genetickú variabilitu. Nová rastlina vzniká zo zygoty, produkovanej splynutím dvoch haploidných gamét. Gamety vznikajú redukčným delením a ich splynutiu predchádza opelenie.
Rodozmena (metagenéza)
Rodozmena (metagenéza) predstavuje životný cyklus charakterizovaný striedaním dvoch geneticky odlišných generácií. Fúziou haploidných gamét vzniká diploidná zygota, čím sa cyklus neustále opakuje a generácie sa striedajú.
- Gametofyt (pohlavná generácia) - tvoria ho bunky s jedným súborom chromozómov (haploidný počet, n). Vzniká mitotickým delením zo spóry. Neskôr vytvára pohlavné orgány (gametangiá), ktoré mitózou produkujú pohlavné bunky (gaméty): plemenníčky (anterídiá) tvoria samčie gaméty (často bičíkaté spermatozoidy vyžadujúce vodu) a zárodočníky (archegóniá) tvoria samičiu bunku (oosféra).
- Sporofyt (nepohlavná generácia) - disponuje homologickými chromozómami (diploidný počet, 2n). Vzniká mitotickým delením zygoty.
Prechod rastlín na súš znamenal evolučný tlak na ochranu citlivých fáz. Vývoj smeroval k posilneniu diploidného sporofytu a redukcii haploidného gametofytu. Prvé suchozemské rastliny, ako ryniorasty, mali ešte primitívnejšiu rovnakotvarú rodozmenu.
Výraznou evolučnou výnimkou sú machorasty (Bryophyta). Dominuje u nich haploidný gametofyt (zelený mach). Sporofyt (stopka s výtrusnicou) je fyziologicky úplne závislý od gametofytu. U cievnatých výtrusných rastlín už dominuje mohutný diploidný sporofyt. Ich drobný gametofyt (prvorast) je síce zredukovaný, ale žije nezávislým životom. Pri semenných rastlinách je gametofyt extrémne zredukovaný a trvalo ukrytý v materskom sporofyte.
Vedeli ste, že...?
Samičí gametofyt krytosemenných rastlín je zredukovaný len na 7 buniek (zárodočný vak) a samčí gametofyt dokonca len na 2-3 bunky (peľové zrnko).
Opelenie a oplodnenie u krytosemenných rastlín
Opelenie (pollinatio) je prenesenie peľu z tyčinky na bliznu piestika. Ak je peľ prenesený z tyčinky na piestik toho istého kvetu, jedná sa o samoopelenie (autogamia). Ak je peľ prenesený na cudzí kvet, je to cudzoopelenie (allogamia). Špecifickým prípadom samoopelenia je kryptogamia (skrytoopelivosť), pri ktorej dochádza k opeleniu ešte v púčiku, teda ešte pred tým, ako kvet rozkvitne.
Peľové zrná (samčí gametofyt) vznikajú v peľniciach. Ich povrch chráni pevná vonkajšia exina a vnútorná tenká intina. Zrelé peľové zrnko krytosemenných rastlín tvoria tri bunky: jedna vegetatívna bunka (zabezpečuje rast peľovej trubice) a dve samčie neobrvené bunky, takzvané spermácie. Práve tieto spermatické jadrá sa zúčastnia oplodnenia ako samčie pohlavné bunky (gaméty).
Zárodočný miešok (samičí gametofyt) sa vyvíja vo vajíčku semenníka. U krytosemenných rastlín sa zostávajúca spóra mitoticky delí 3-krát, vytvorí sa útvar s ôsmimi haploidnými jadrami. Tento útvar sa označuje ako zrelý zárodočný miešok krytosemenných rastlín. Zrelý zárodočný miešok semenných rastlín je po opelení pripravený na oplodnenie.
Prenos peľu na bliznu označujeme ako opelenie. Podľa pôvodu peľu rozlišujeme samoopelenie (autogamiu) a cudzoopelenie (alogamiu). Rastliny sa samoopeleniu bránia autoinkompatibilitou (biochemický blok) alebo časovým nesúladom dozrievania orgánov.
Peľ môže byť prenesený pomocou vetra - vetroopelivé rastliny, prostredníctvom živočíchov najčastejšie hmyzu (včely), ale aj vtákov (kolibríky) alebo malých cicavcov. U niektorých rastlín môže peľ preniesť voda.
Dômyselný priestorový mechanizmus na podporu cudzoopelenia využíva prvosienka jarná. Jednotlivé rastliny tvoria buď kvety s dlhými čnelkami a krátkymi tyčinkami, alebo presne naopak, čo priamo núti hmyz prenášať peľ výhradne medzi rôznymi jedincami.
Ak na bliznu dopadne správny peľ, prítomný vápnik, bór a fytohormóny ho okamžite stimulujú ku klíčeniu. Proces dvojitého oplodnenia je unikátom krytosemenných rastlín. Po vniknutí trubice do vajíčka prebehnú dve splynutia súčasne:
- prvá spermácia splynie s oosférou, čím vzniká diploidná zygota (budúce embryo).
- druhá spermácia splynie s centrálnou bunkou, čím vzniká bunka s triploidným jadrom. Z nej sa sformuje zásobný triploidný endosperm vyživujúci embryo.
U nahosemenných rastlín splynie spermatické jadro s vajcovou bunkou a vytvorí sa zygóta. Zásobné pletivo (endosperm) je haploidné a je vytvorené pred oplodnením. U krytosemenných rastlín splynie jedno spermatické jadro s vajcovou bunkou - vzniká zygóta. Druhé spermatické jadro sa spojí s centrálnym jadrom zárodočného mieška a postupne sa z neho delením vytvorí triploidný (3n) endosperm.
Splynutím samčej a samičej gaméty nastalo oplodnenie.
Strata opeľovačov. Aké následky môže ľudstvo očakávať?
Vývoj semena a plodu
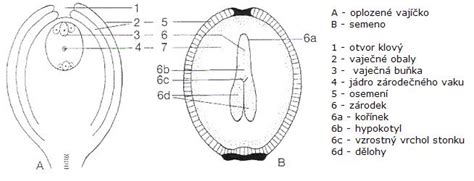
Z oplodneného vajíčka sa vyvíja semeno chránené pevným osemením (testa), ktoré vzniklo z pôvodných vajíčkových obalov. Pre úspešný vývin a následné klíčenie sa v semene, predovšetkým v endosperme, koncentrujú kľúčové živiny a veľké množstvo fosforu. Semeno následne stratí väčšinu vody (jej obsah klesne na 5-20 %) a prechádza do ochranného stavu spánku (dormancie).
Výnimočne môže semeno vzniknúť aj úplne bez oplodnenia. V zoológii sa vývin z neoplodneného vajíčka nazýva partenogenéza, no v botanike tento jav produkcie klonálneho semena označujeme ako apomixia.
Krytosemenné rastliny (iné názvy: krytosemenné, angiospermy, kvitnúce rastliny, kvetnaté rastliny; lat. Angiospermae, Angiospermatophyta) alebo magnóliorasty (Magnoliophyta) sú veľký taxón rastlín. Sú to semenné rastliny, ktoré majú semená ukryté v plode. Odlišná je klasifikácia podľa systému APG IV, pretože sú v nej dvojklíčnolistové rastliny rozdelené na viacero samostatných taxónov, medzi ktoré je kvázi uprostred "vnorený" taxón jednoklíčnolistové rastliny.
V semene krytosemennej rastliny je uložené embryo. Za vhodných podmienok začne semeno klíčiť a vyrastá z neho diploidný sporofyt. V prvom štádiu vývoja klíčiaceho semena sa objavujú klíčne listy, ktorých počet býva niekedy použitý ako základ klasifikácie krytosemenných rastlín (na jedno- a dvojklíčnolistové rastliny).
V čase pohlavnej dospelosti sa na rastline, sporofyte, utvoria kvety. Zrastením viacerých samičích pohlavných orgánov - výtrusníc - vzniká vajíčko (prostredná výtrusnica) a vajíčkové obaly (ostatné výtrusnice). Obaly vajíčka - integumenty - sú dva, vzácnejšie jeden, a obrastajú celé vajíčko. Na vrchole však zostáva nezrastený otvor - peľový vchod. Vajíčko je chránené plodolistami, ktoré niekedy zrastajú a vytvárajú piestik. V jednom piestiku sa môže nachádzať aj viac vajíčok. Vajíčka sú spojené s tzv. placentou vajíčkovou šnúrou.
V mnohobunkovom vajíčku vzniká jedna veľká bunka - materská bunka zárodočného mieška. Tá vstupuje do redukčného (meiotického) delenia, po ktorom vzniknú 4 haploidné bunky. Tri z nich zaniknú, štvrtá, sa nazýva jednobunkový mladý zárodočný miešok. V ňom prebehnú 3 mitotické delenia a vznikne tak 8-jadrový 7-bunkový útvar. V jeho strede vzniká diploidné centrálne jadro, k hornému koncu útvaru sa premiestni vajcová bunka - oosféra. Ostatné bunky sa rozdeľujú na synergidy a antipódy.
Samčie pohlavné orgány sú tyčinky. V tyčinkách vznikajú peľové komôrky a v nich peľové zrná. V čase zrelosti sa peľové zrná skladajú z jednej vegetatívnej a jednej generatívnej bunky, alebo z jednej vegetatívnej a dvoch generatívnych buniek. Peľové zrnko sa musí nejakým spôsobom dostať na bliznu piestika (za pomoci vetra, živočíchov...). Tento proces na nazýva opelenie.
Na blizne začne peľové zrnko klíčiť a vzniká peľové vrecúško. To preniká až k vajíčku. Jedna generatívna bunka oplodní vajcovú bunku, z nej vznikne zygota a postupným mitotickým delením zárodok budúcej rastliny (embryo). Druhá generatívna bunka oplodní centrálne jadro, ktoré je však už pred oplodnením diplodiné, preto oplodnením vzniká triploidná bunka, z ktorej sa utvorí triploidný tzv. sekundárny endosperm.
Vajíčko sa postupne zmení na semeno a plodolisty na oplodie. Antipódy a synergidy zaniknú. Blizna a čnelka a nepodieľajú na vzniku plodu a väčšinou tiež zaniknú.
Strata opeľovačov. Aké následky môže ľudstvo očakávať?
tags: #embryo #krytosemennych #rastlin